
近日,国际著名期刊Nature Ecology & Evolution(IF:16.8),发表了一篇题为“ Carbohydrate complexity limits microbial growth and reduces the sensitivity of human gut communities to perturbations”的研究论文。论文通过建立模型分析了膳食纤维的化学复杂性对微生物相互作用和对扰动的反应的影响。研究发现,纤维的复杂性限制了微生物的生长和利用纤维的物种数量。此外,在纤维存在下,负相互作用减少,群落聚集能力和抵抗入侵的能力增强。最大或最小化居民和入侵物种之间的碳水化合物竞争可以增强抵抗入侵的能力。总之,纤维的化学复杂性对微生物相互作用网络有定量影响,这对我们理解膳食和微生物的关系很重要。这些生态学原理可用于指导饮食和细菌治疗干预措施的设计,以提高人类肠道微生物群对环境扰动的适应力。

图1 文章发表信息
研究背景
人类肠道微生物群是一个复杂的微生物生态系统,在人类生理、营养、行为和疾病状态中发挥着重要作用。人类肠道微生物群的动态和功能活动是由环境变量(如饮食因素)和群落组成成员和宿主之间的生物相互作用的复杂组合不断形成的。膳食纤维的摄入极大地影响了肠道群落的动态和功能,并与健康益处相关。然而,我们缺乏预测肠道群落动态随膳食纤维变化而变化的能力。因此,有必要详细和定量地了解化学多样性的纤维如何塑造人类肠道群落的生物相互作用和社区水平的反应。
研究思路
测序策略:二代扩增子测序
测序平台:Illumina NovaSeq
研究结论
(1)建立人类肠道菌群群落与计算模型,剖析化学复杂性和糖类组成不同的纤维如何影响菌群组装以及菌群对扰动的反应;
(2)纤维的化学复杂程度会限制菌群的生长和利用该营养物质的菌种数量;
(3)与组成糖相比,纤维的存在减少菌种间竞争;
(4)碳水化合物的化学复杂性增强群落组成的可重复性,提高群落对入侵的抵抗力;
(5)最大化或最小化常驻和入侵菌群间对碳水化合物的竞争,可以增强对入侵的抵抗力。
研究结果展示
1、物种生长是碳水化合物复杂性的函数
为了了解膳食纤维对群落动态和系统水平行为的影响,研究者构建了一个多样化的合成人体肠道群落来代表一个健康的人体肠道微生物群。一个确定的群落可以表征每个物种对单一碳水化合物的生长反应。10个成员的群落(Comm10)跨越了人类肠道的关键代谢生态位,包括拟杆菌门(Bacteroides thetaiotaomicron, BT; Bacteroides vulgatus, BV; and Para bacteroides johnsonii, PJ),具有灵活代谢能力的物种,包括氨基酸发酵(Eggerthella lenta, EL; and Clostridium scindens, CS)、硫酸盐还原剂(Desulfovibrio piger, DP)和H2消耗乙酰原(Blautia hydrogenotrophica, BH)。研究者描述了个体物种在24小时内在单一碳水化合物浓度范围内的生长情况。为了定量评估每个物种的碳水化合物生长响应曲线,我们确定了单栽培生长曲线下的总面积(积分OD(intOD)或总生长;方法)。intOD受到生长响应的所有特征的影响,包括滞后阶段、增长率和承载能力。使用这个指标将每个物种分类为一个代谢生态位。碳水化合物反应者被定义为在没有任何碳水化合物时,有特定碳水化合物的总生长显著增加的物种(在至少两种浓度的碳水化合物中,非配对t检验P<0.05)。碳水化合物无反应者被定义为相对于无碳水化合物对照,在给定碳水化合物存在下总生长没有显著增加的物种(扩展数据图1a,在给定碳水化合物浓度下的非配对t检验P<0.05)。因此,每一种碳水化合物类型都有一组独特的应答者和无反应物种。
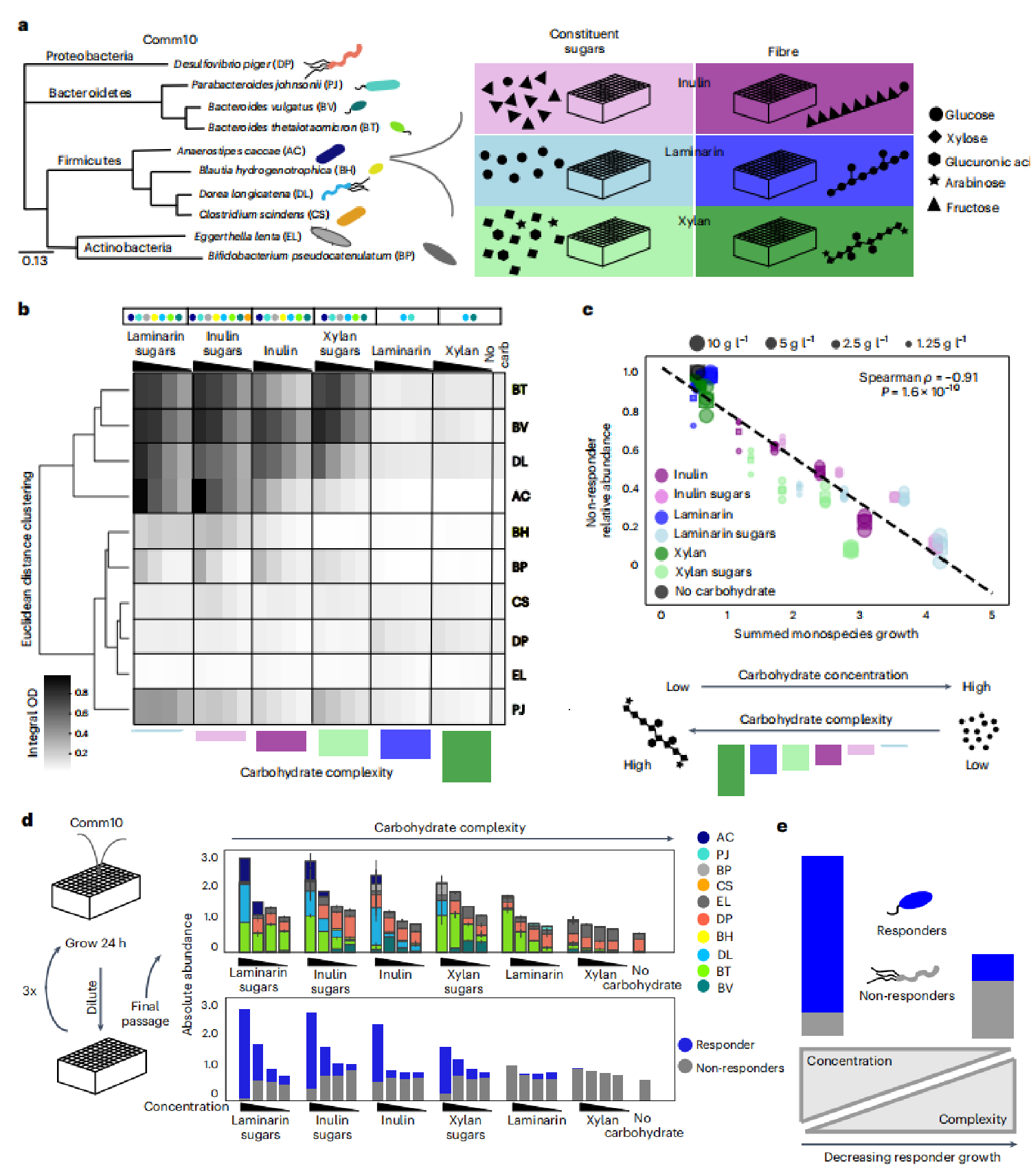
图2 复杂的碳水化合物限制了单个物种的生长,并增加了群落内碳水化合物无反应者的丰度
2、纤维减少了微生物的竞争
微生物群落的组装是由一个复杂的相互作用网络所决定的。为了确定物种间相互作用对群落聚集的贡献,使用了一个假设没有物种间相互作用的逻辑生长模型(即零模型)来预测每个培养基中的物种丰度。这个零模型在存在大多数碳水化合物时不能很好地预测群落组成。为了解释物种间的相互作用,设计了包含有反应者和无反应者的独特组合的亚群落,并在单个碳水化合物存在下的24小时内确定了群落组合的特征。为了解读在不同碳水化合物存在下,物种间的相互作用影响了群落的组装,我们使用了gLV动态生态模型,该模型捕获了物种生长作为每个物种内在生长速率的函数,以及与所有组成群落成员的成对相互作用。gLV模型的参数可以在物种绝对丰度测量的基础上进行优化技术估计。使用经验贝叶斯框架来获得gLV参数的最大后验(MAP)估计值。该方法提供了超参数的MAP估计,如测量方差和先验参数方差,这决定了正则化的程度。通过交叉验证,每个模型与保留的数据(PearsonR=0.75–0.93,P<0.0001)具有良好的一致性,海带素和海带素的糖含量最低。纤维的存在促进了物种间负相互作用和正相互作用的百分比之间的平衡,而在糖存在时,负相互作用超过了正相互作用。使用Wald检验来排除没有受到我们的训练数据约束的参数,以进一步获得对不同条件下的趋势的信心。通过考虑充分约束的种间相互作用系数,纤维还促进了正相互作用和负相互作用比例之间的平衡。因此,纤维中糖苷键的存在减少了微生物的竞争,并促进了网络中更相似比例的正相互和负相互作用。
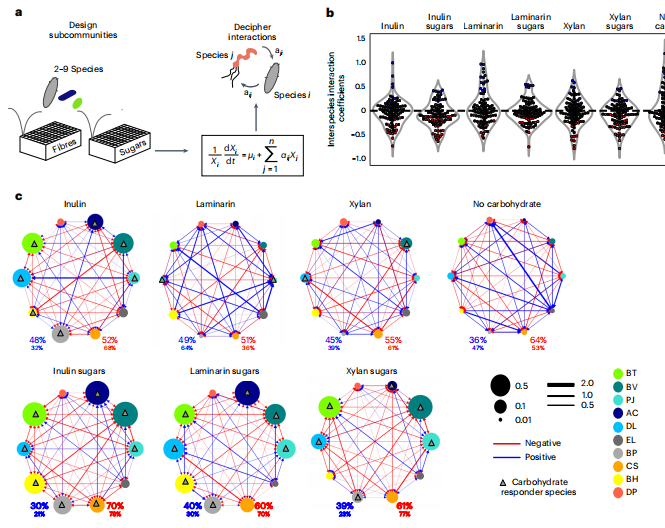
图3 复杂的碳水化合物通过减少反应者之间的竞争,促进物种间积极和消极相互作用比例的平衡
3、代谢生态位的占据决定了相互作用的标志
有反应者和无反应的数量因碳水化合物类型而异。因此,我们考虑了所有有反应者和无反应者组合之间的四类相互作用。我们通过在一个给定的类别中可能的交互作用的总数来标准化推断的交互作用的数量。例如,对于木聚糖糖中的应答-应答者组,有6种应答者物种,因此有30种可能的应答-应答者相互作用。从有反应者到无反应者的积极相互作用与多糖分解产物或代谢副产物的释放机制相一致。然而,相对于组成糖,在纤维存在的情况下,正反应者对无反应者相互作用的比例更低。相反,在所有碳水化合物类型中,无反应者之间更频繁地观察到积极的相互作用。t无反应者相互作用的符号在所有碳水化合物类型中都是一致的。因此,纤维存在时正相互作用频率的增加是由于群落中无反应物种的丰度的增加。同样,在所有碳水化合物培养基中,反应者对之间的相互作用通常是负的而不是正的。这意味着,诸如资源竞争、有毒代谢副产品或抗菌素释放等负面相互作用可以介导应答物种之间的相互作用,而不管碳水化合物类型如何。总之,有应答者或非应答者代谢生态位的占据是影响物种间相互作用分布的主要因素。
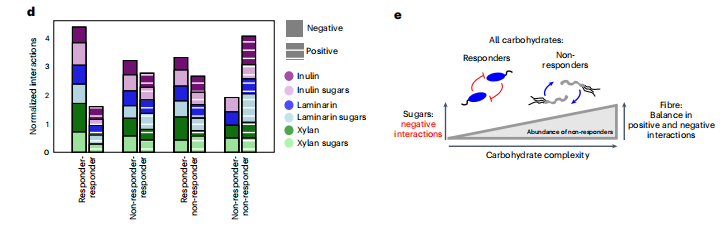
图4 在碳水化合物应答者或非应答者的代谢生态位中,物种间相互作用系数的归一化数
4、生长限制降低了对入侵的敏感性
在人类的肠道中,微生物经常从外部环境中被引入。然而,由于健康的肠道微生物群提供的定植抗性,这些微生物在入侵后的持续生存能力已被证明是高度可变的。资源竞争是一种被提出的定殖抗性机制。因此,研究了碳水化合物复杂性对居民群落对入侵的抵抗力以及入侵者在群落中生存的能力的作用。入侵者(16种不同的肠道物种)是根据之前的工作中广泛的特征特征选择的。在复杂纤维层粘连素或木聚糖的存在下,一小部分个体入侵物种的生长得到了增强。此外,碳水化合物的复杂性和个体入侵物种的总生长呈负相关,与Comm10物种的生长趋势一致。为了调查对入侵的反应,居民社区(Comm10)在存在或不存在个体碳水化合物的情况下生长,并传代两次,以允许居民社区聚集。在第二代,引入了26种肠道物种(包括所有Comm10种)的混合物,其丰度低于居民群落或等量的无菌培养基。在入侵过程中,居民群落中的物种被重新引入,以模拟暴露于环境水库,这已被证明塑造了群落对扰动的反应。在入侵扰动后,所有群落被传代两次,以在较长的时间尺度内跟随群落聚集。我们通过计算最终时间点存在和不存在入侵时的居民群落组成的欧氏距离来评估群落对入侵的敏感性(方法)。
居民群落对入侵的敏感性是一个与健康相关的指标,因为病原体等入侵者可以改变肠道的代谢景观,导致居民物种丰度的持续变化并延长。碳水化合物的复杂性与入侵引起的居民群落组成的变化呈很强的负相关关系。假设由于资源竞争,与常驻物种共享一个碳水化合物利用生态位的入侵物种会对常驻物种的生长产生负面影响。可以证实这一观点的是,在存在入侵的情况下,响应物种的丰度比无响应物种的丰度变化更大。Comm10在存在糖的情况下的比例高于纤维,这与这些条件下入侵和未入侵群落组成之间的欧氏距离一致。Comm10种的单种生长总量与入侵后居民群落组成的变化呈正相关。这表明,物种之间的竞争相互作用和生长对环境的耦合修饰驱动了入侵过程中居民群落组成的破坏程度。我们评估了抗入侵能力作为碳水化合物浓度的函数,以确定通过降低糖浓度来限制生长是否可以反映对入侵的“纤维样”反应。居民群落(Comm10)在一定的碳水化合物浓度范围内生长,并被如前所述的26个成员的群落入侵。当考虑所有碳水化合物浓度时,入侵后居民群落组成的变化与单种生长总量没有显著相关性。这表明,在低碳水化合物浓度下,未知的相互作用或未知的欧氏距离的变化,影响了群落对入侵的抗性。然而,在所有媒介的碳水化合物浓度降低时,存在入侵和没有入侵的居民群落的欧氏距离显著降低(双侧非配对t检验p<0.05)。
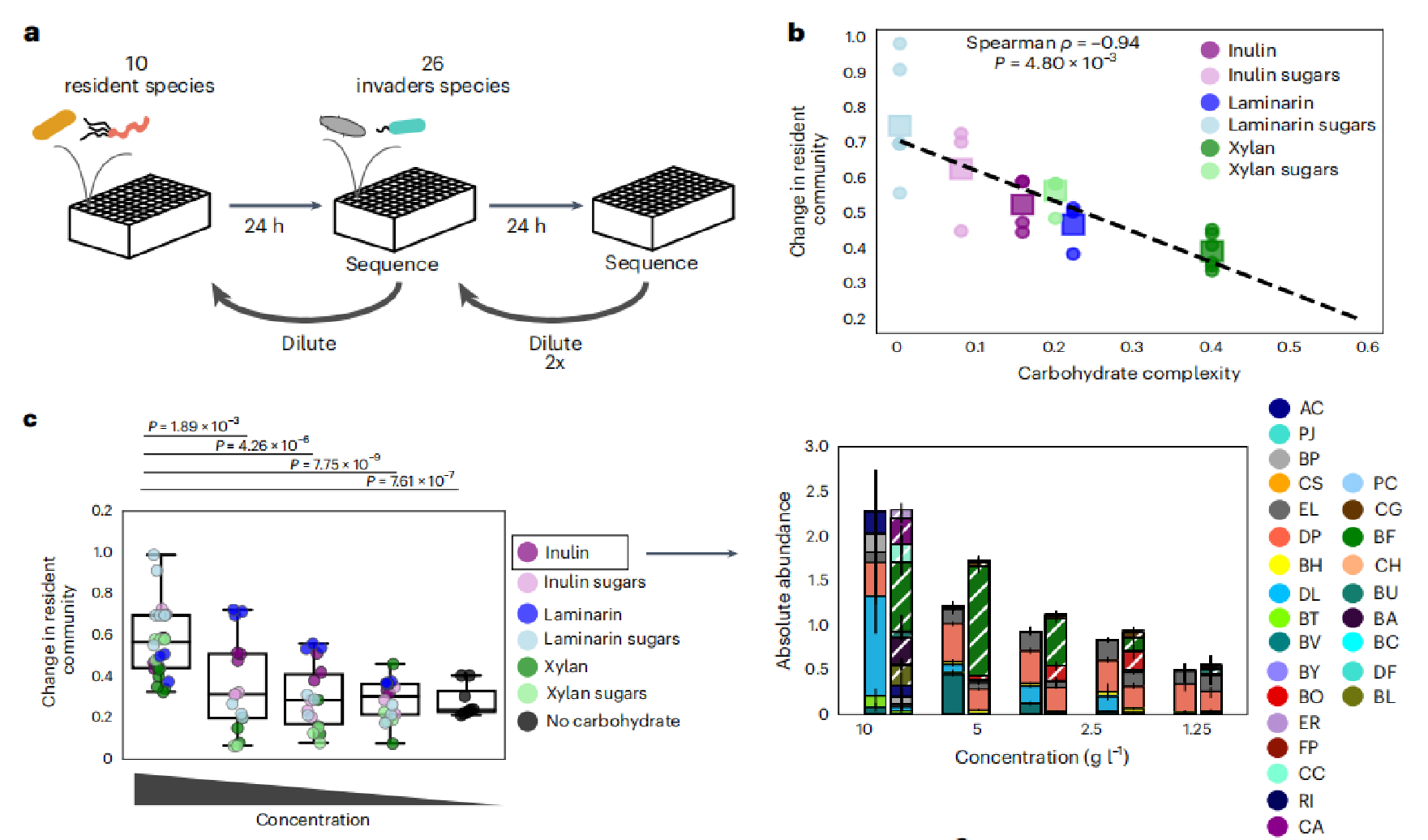
图5 碳水化合物的复杂性、碳水化合物浓度的降低和生态竞争的程度影响着群落对入侵的抵抗力
 微信公众号
微信公众号


 027-87224696
|
027-87224696
| marketing@frasergen.com
|
marketing@frasergen.com
|
