
近日,华中科技大学栗茂腾教授团队在Plant Communications(IF:10.5)在线发表了题为 “3D genome structural variations play important roles in regulating seed oil content in Brassica napus”的研究成果,为从基因组三维结构角度理解SOC的分子调控机制提供了新的视角。首先,作者利用Hi-C、ATAC-seq和RNA-seq多组学技术,比较了高油材料N53-2(含油量超过50%)和低油材料Ken-C8(含油量约40%)中基因组三维结构、染色质开放性和基因表达的差异,并发现N53-2和Ken-C8之间存在大量DARs和DEGs,这些DARs和DEGs倾向于富集在SOC相关QTL/AGRs区域中,并且SOC相关基因的表达水平与QTL/AGRs区域中的基因组结构变异密切相关。其次,作者通过构建N53-2和Ken-C8杂交产生的KN-DH群体并进行精细定位,鉴定出了A09染色体QTL/AGRs区域中存在结构变异的候选基因BnaA09g48250D,该基因被注释为乙酰辅酶A羧化酶2(CAC2),是脂肪酸合成过程中的关键酶。最后,作者通过基因功能验证实验,表明BnaA09g48250D的过表达和敲除分别导致转基因株系的SOC显著增加(4.90% ~ 9.46%)和减少(3.03% ~ 6.75%)

图1 文章信息
研究思路
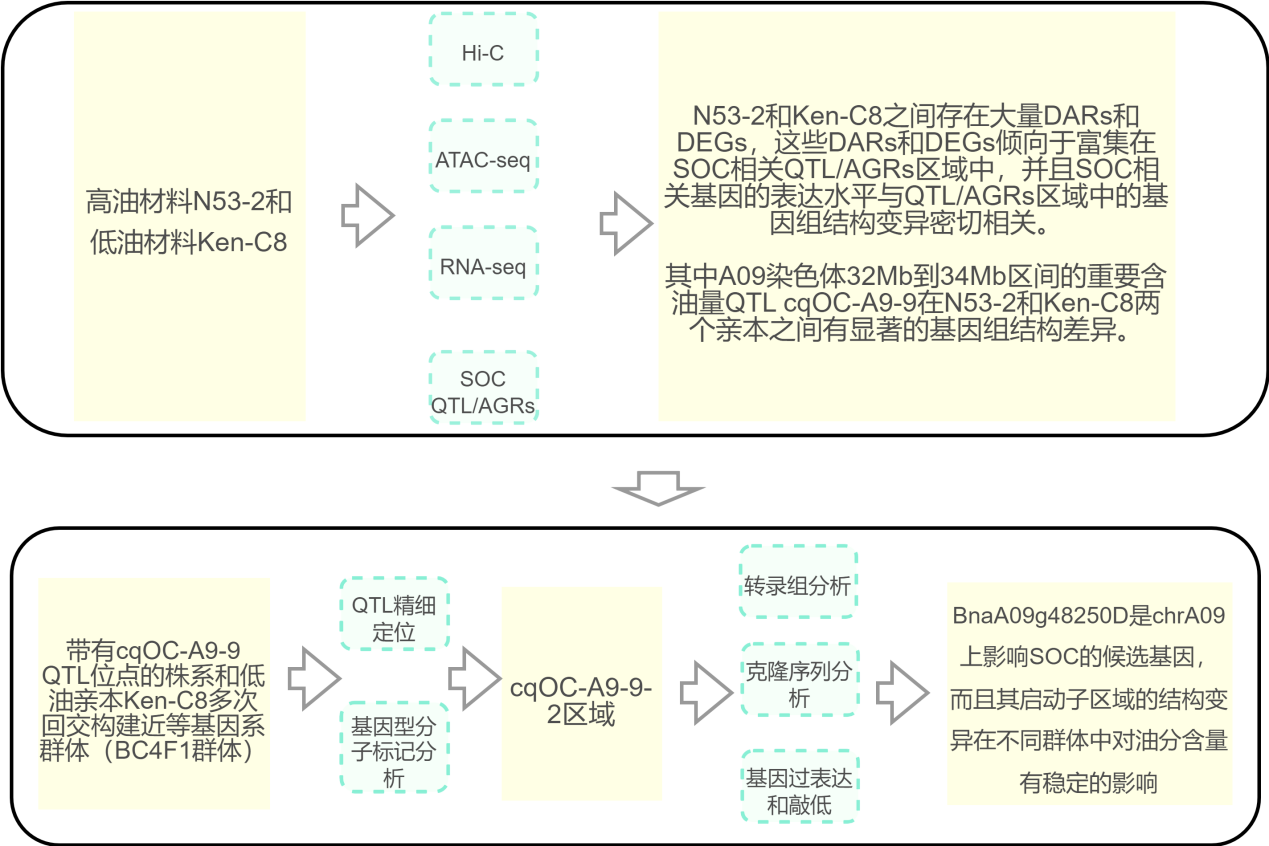
研究结果
1、N53-2和Ken-C8的基因组三维结构和基因组结构变异
为了探索甘蓝型油菜种子基因组三维结构与油分含量(SOC)之间的关系,该研究对高油材料N53-2(含油量超过50%)和低油材料Ken-C8(含油量40%左右)的种子进行Hi-C实验,利用Juicer软件分析了染色质相互作用,并重构了3D基因组结构。首先,该研究发现N53-2中有159675个染色体内相互作用和1787062个染色体间相互作用(图2A),而Ken-C8中有163267个染色体内相互作用和1765908个染色体间相互作用(图2B)。其次,该研究发现N53-2和Ken-C8的3D基因组结构存在局部结构差异。例如,N53-2中端粒(蓝色点)主要分布在外围区域,而Ken-C8中端粒浸没在内部区域(图2C-1F),这表明Ken-C8中3D基因组结构比N53-2中更紧凑。
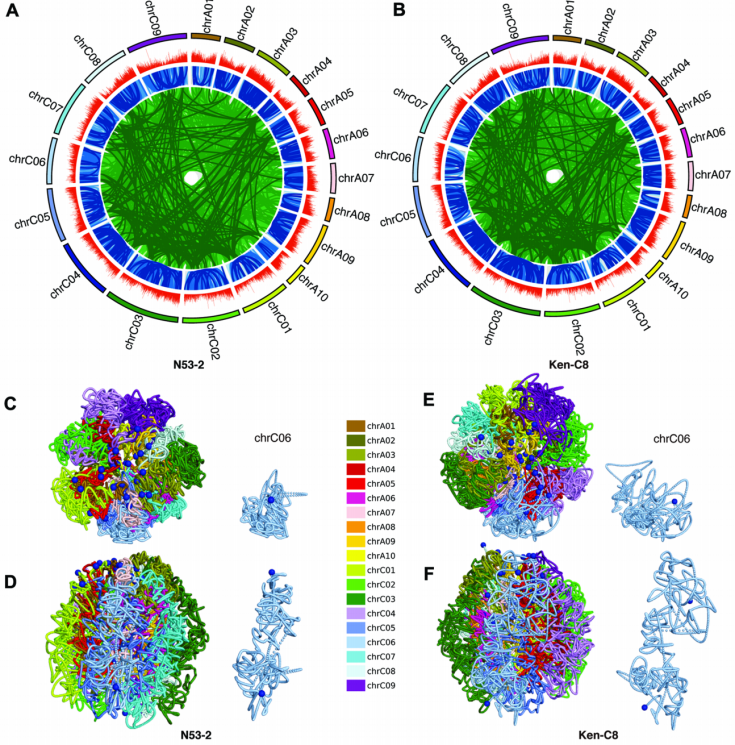
图2 N53-2和Ken-C8的相互作用分析和3D基因组结构
此外,该研究发现无论是在N53-2还是在Ken-C8中,A02和C02染色体之间有最高的相互作用频率,A亚基因组内部的相互作用频率远大于C亚基因组内部和A与C亚基因组之间的,A与C亚基因组之间的同源区域的相互作用频率显著高于非同源区域的。同时,A亚基因组的整体基因表达水平高于C亚基因组。A与C亚基因组之间都有很高的同源性。另外,在多条染色体上存在很多可能形成KNOT结构和KEEs(KNOT-engaged elements,异染色质区域内一些远距离互作网络形成的结构)的强染色质相互作用。这些结果表明,油菜种子基因组三维结构具有复杂的特征和差异,可能与SOC有关。
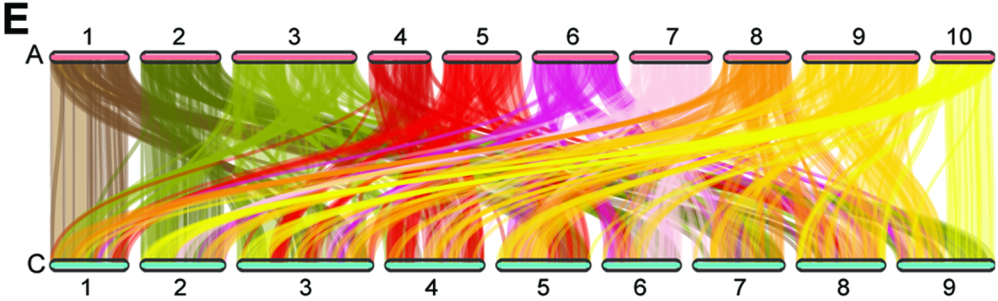
图3 N53-2 A亚基因组和C亚基因组的共线性分析
为了进一步分析N53-2和Ken-C8中染色质的功能状态,该研究结合前期得到的数量性状位点/关联区域(QTL/AGRs),对compartment和TADs的特征进一步探索,首先,N53-2和Ken-C8分别有556个和648个compartment,以及909和907个TADs。其次,QTL/AGRs中A compartment的比例比全基因组中的高,而B compartment并非如此。此外,A compartment的基因密度比B compartment的高。这些结果表明,QTL/AGRs区域中的染色质比其他基因组区域更活跃。同时,在TAD结构中,N53-2中的KEEs信号比Ken-C8中的强,表明N53-2中的染色质更容易形成KNOT结构(图4 E-F的蓝色箭头)。对TAD边界区域进行的motif分析显示,MYB46和WRKY28转录因子(TF)可能在油菜基因组边界区域发挥重要的调控作用。
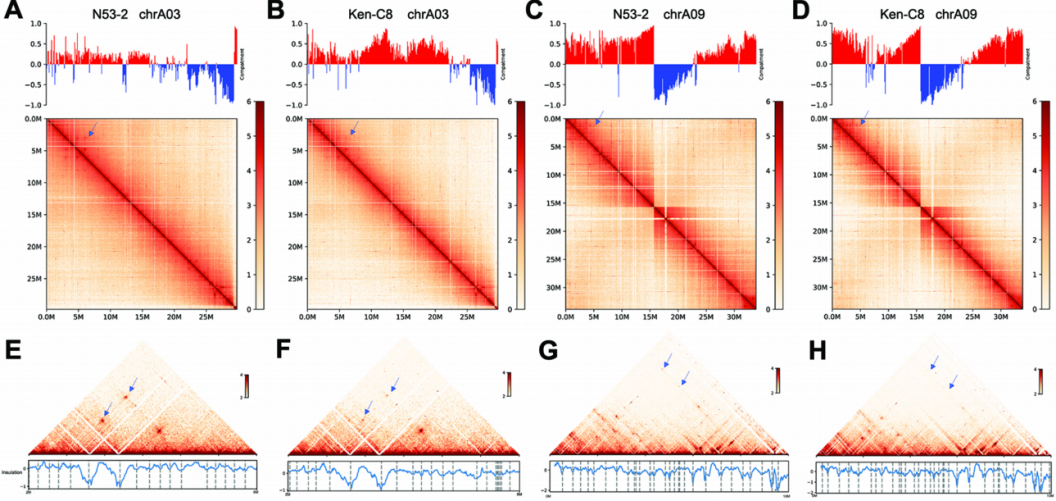
图4 Compartment和TAD分析
2、QTL/AGRs区域的染色质开放性和基因表达
该研究对高油材料N53-2和低油材料Ken-C8进行ATAC-seq和RNA-seq实验,识别了差异开放性区域(DARs)和差异表达基因(DEGs)。首先,该研究发现N53-2和Ken-C8之间存在121345个DARs,其中N53-2相对于Ken-C8有63722个开放性增强的区域和57623个开放性降低的区域。其次,N53-2相对于Ken-C8有7473个上调基因和6247个下调基因。这些差异表达基因与SOC相关的KEGG通路有关,如“亚油酸代谢”、“甘油磷脂代谢”、“脂肪酸生物合成”和“脂肪酸代谢”。此外,该研究结合前期SOC相关的QTL/AGRs数据进行比较分析。结果显示,N53-2和Ken-C8之间的DARs和DEGs富集在QTL/AGRs主要分布的染色体上(图5)。同时,进一步分析DARs在QTL/AGRs区域内外的分布和密度,发现DARs倾向于QTL/AGRs区域内。与此类似,N53-2和Ken-C8之间的DEGs,尤其是上调DEGs也倾向于QTL/AGRs区域内。这些结果表明,在N53-2和Ken-C8中,QTL/AGRs区域中的染色质开放性和基因表达与SOC有关。
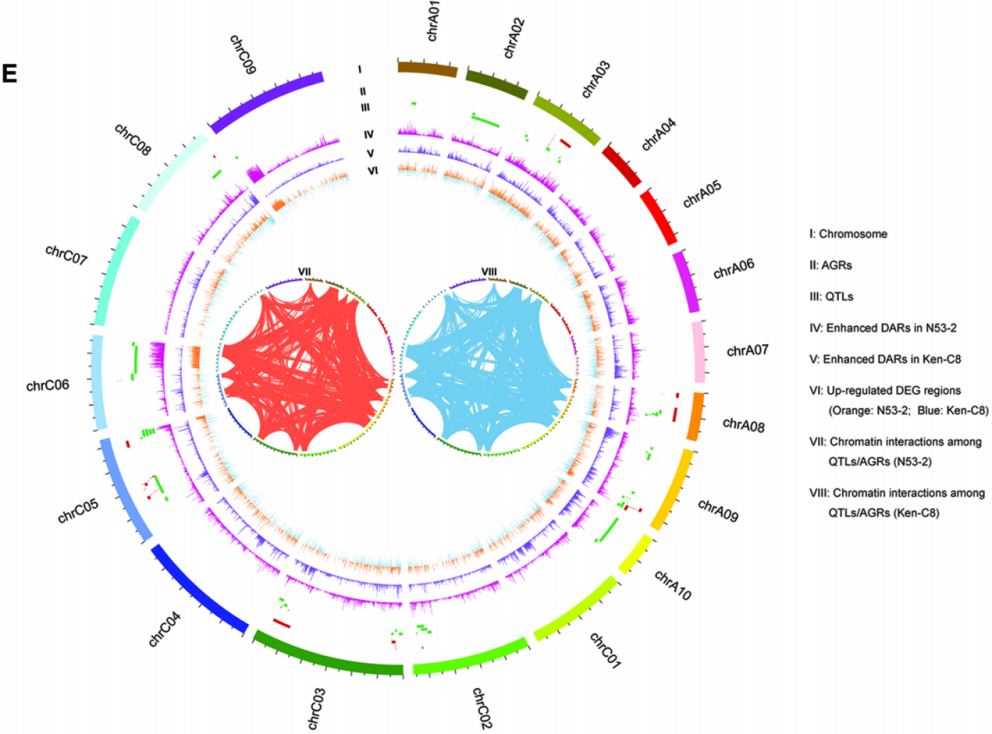
图5 N53-2和Ken-C8的DARs、DEGs和QTL/AGRs
3、基因组结构变异影响QTL/AGRs区域的基因表达
该研究对N53-2和Ken-C8进行多组学整合分析。研究发现,一些QTL/AGRs区域在N53-2和Ken-C8之间存在显著的结构变异(图6)。其次,该研究分析了位于QTL/AGRs区域内DARs相关联基因表达水平,并发现N53-2中开放性增强DARs相关联基因的整体表达水平也显著更高。由于DARs反映了N53-2和Ken-C8之间染色质开放性的差异,因此基因组结构变异可能与QTL/AGRs区域内基因表达水平有关。
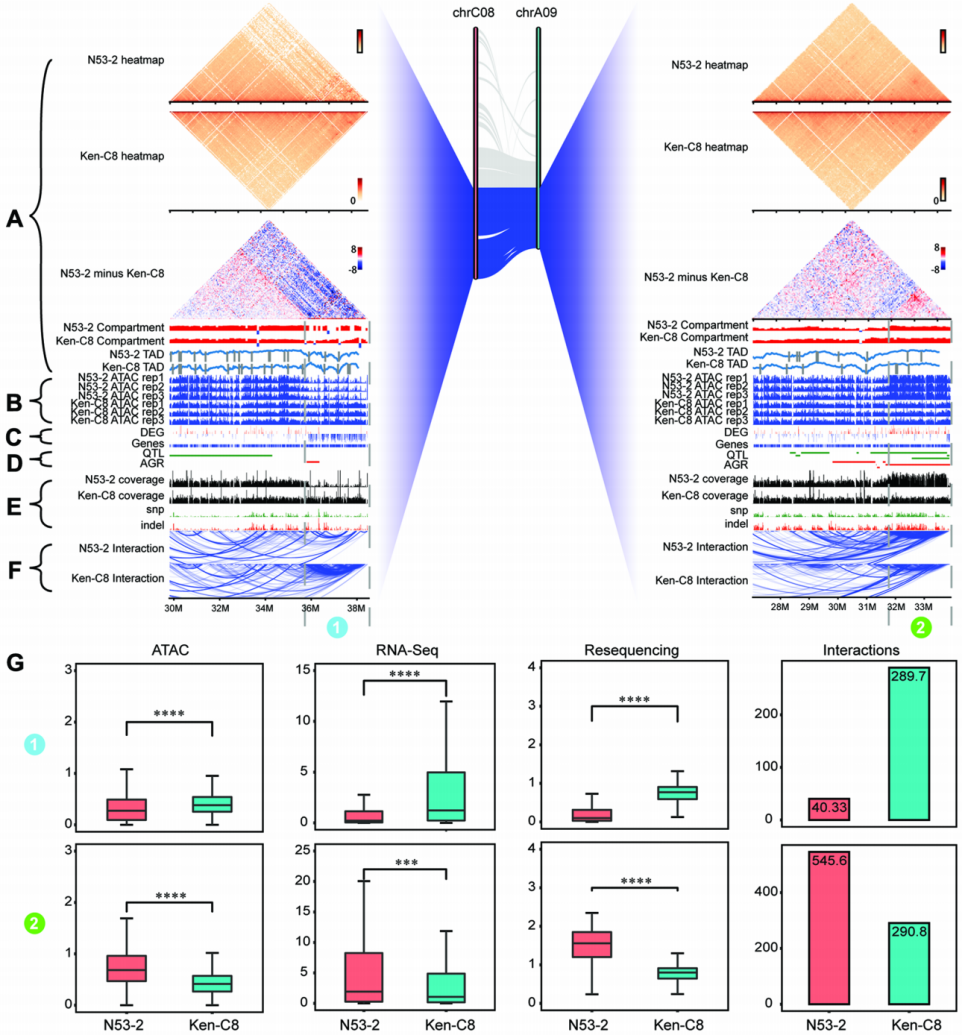
图6 N53-2和Ken-C8中chrC08(左)和chrA09(右)的多组学分析(Hi-C、ATAC-seq、RNA-seq、QTL/AGR、重测序和染色质相互作用)
此外,一些重要的DEGs(包括转录因子和参与脂肪酸生物合成和三酰甘油形成的基因)主要分布在QTL/AGRs富集区域,并且在N53-2中相对于Ken-C8上调。该研究还探索了转录因子与参与脂肪酸合成通路的基因之间的相互作用,共鉴定出50个相互作用位点,并发现N53-2和Ken-C8之间存在相互作用模式的差异。例如N53-2中LEC2、FUS3和ABI3与chrA10上的BnaA10g01090D(GPAT2)发生相互作用。已有研究表明,ABI3通过参与植物油合成的相互作用网络来激活油积累。然而,在SOC低的Ken-C8中,这种相互作用被打断了。
4、chrA09 QTL/ARGs区域的精细定位和候选基因的功能验证
该研究通过多组学分析,发现了一个位于A09染色体32Mb到34Mb区间的重要含油量QTL cqOC-A9-9,该区域在N53-2和Ken-C8两个亲本之间有显著的基因组结构差异。该研究在筛选带有该QTL位点DH株系的基础上,通过和低油亲本Ken-C8多次回交构建了包含2820个单株的BC4F1 NILs(近等基因系)群体,并根据Ken-C8和N53-2的重测序结果开发Indels分子标记,对cqOC-A9-9进行了精细定位,将其分为两个区域cqOC-A9-9-1和cqOC-A9-9-2,其中,cqOC-A9-9-2覆盖了一个300Kb的区域,包含了62个注释基因,被选为进一步研究的对象。
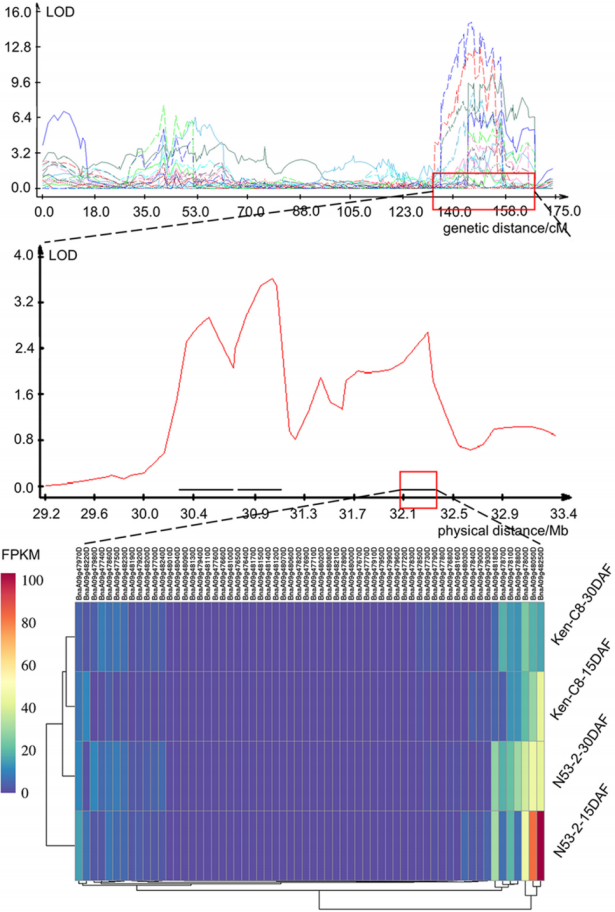
图7 cqOC-A9-9的精细定位
根据转录组分析,该研究发现cqOC-A9-9-2内有19个DEGs,其中BnaA09g48250D编码异源乙酰辅酶A羧化酶(htACCase)的生物素羧化酶亚基,是唯一一个与油脂合成相关的基因。该研究通过克隆测序分析发现BnaA09g48250D编码区没有差异,但Ken-C8中其启动子区有一个6313bp的插入,并且该区域在Ken-C8的染色质相互作用相对较少。随后通过过表达和CRISPR/Cas9敲除的方法,证实了这个基因对SOC的调控作用:过表达BnaA09g48250D的转基因株系比野生型显著增加SOC,而敲除BnaA09g48250D的转基因株系比野生型有显著降低SOC。
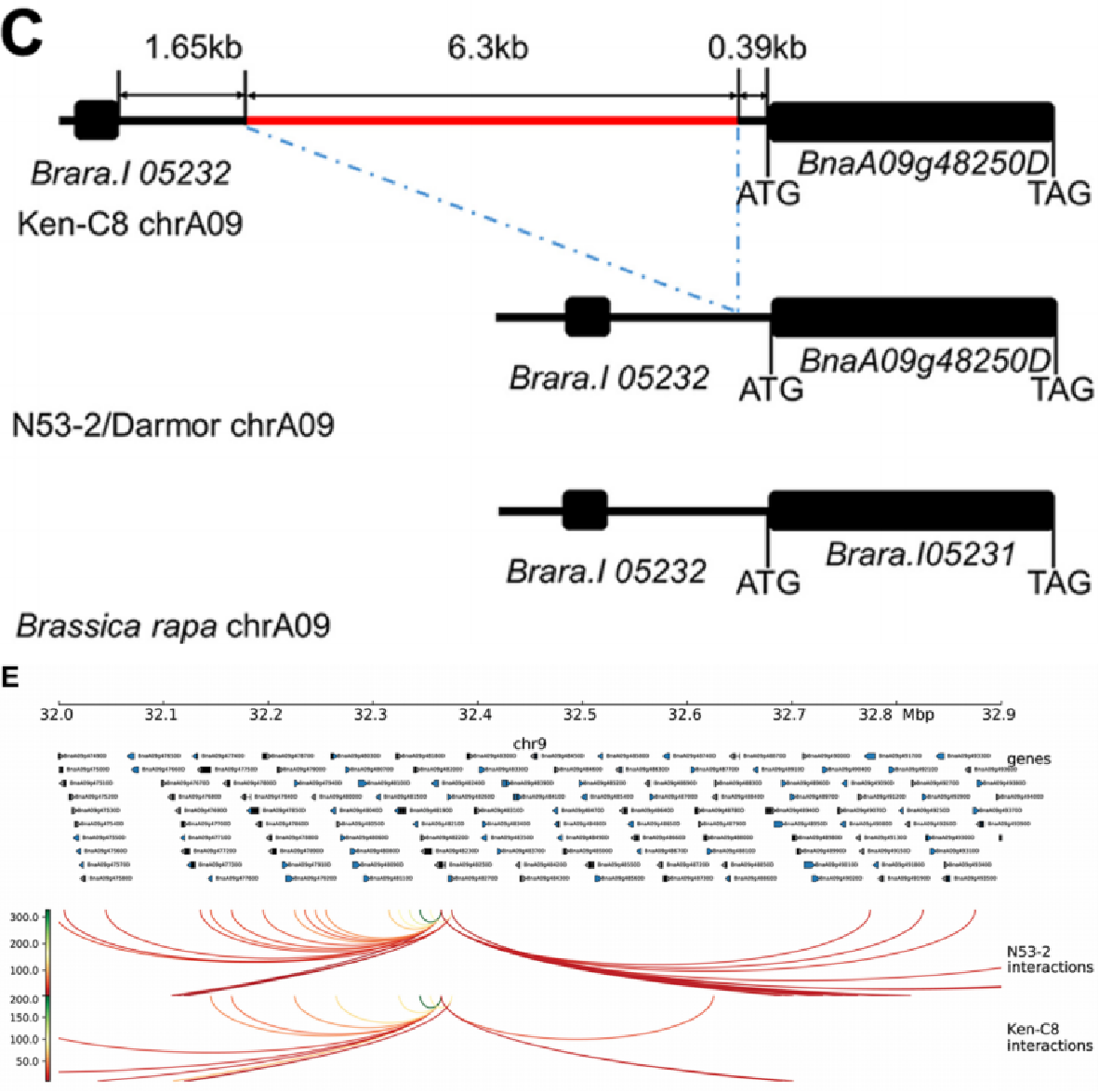
图8 Ken-C8和N53-2中BnaA09g48250D的序列分析(上)及其与相邻区域相互作用的信号(下)
最后,该研究通过KN DH群体在不同环境和年份下的SOC数据,以及一个包含chrA09上与SOC QTL热点共定位的DAR的近等基因系,证实了BnaA09g48250D是chrA09上影响SOC的候选基因,而且其启动子区域的结构变异在不同群体中对SOC有稳定的影响。
小 结
该研究通过Hi-C揭示了甘蓝型油菜基因组3D结构特征,并结合ATAC-seq、RNA-seq、QTL/AGRs等方法探讨了基因组结构变异在甘蓝型油菜SOC调控中的作用。该研究证实了BnaA09g48250D基因是chrA09上影响SOC的候选基因,而且其启动子区域的结构变异在不同群体中对SOC有稳定的影响,这为油菜种子油分含量调控网络及其遗传改良策略提供了新的线索和依据。
华中科技大学生命科学与技术学院的栗茂腾教授为本研究的通讯作者,张礼斌副教授、李怀鑫博士生及澳门·永利集团3044的刘琳博士为共同第一作者,何坚杰和闫书祥博士生、朝红波、尹永泰和赵卫国博士也参与了此项研究。该研究得到国家重点研发计划项目“油菜和花生重要基因资源挖掘与利用”(2022YFD1200402)和国家自然科学基金面上项目(32272067 和32072098)的资助。
 微信公众号
微信公众号


 027-87224696
|
027-87224696
| marketing@frasergen.com
|
marketing@frasergen.com
|
