
生物多样性是地球生命的基础,保护生物多样性、促进可持续发展是全人类共同的责任和义务,同时10月11日-15日在昆明举行的CBD COP15会议也明确指出要推动制定“2020年后全球生物多样性框架”。而保护生物多样性,最重要的一项工作就是保护珍稀濒危物种,减少物种灭绝事件的发生。
科学保护及利用濒危物种资源的前提是了解濒危物种的遗传组成及演化规律,通过构建濒危物种的参考基因组,我们一方面可以解析濒危物种狭小生态位的适应性基础,另一方面我们可以从濒危物种中寻找新的遗传变异,来为重要物种农艺性状的改良提供有价值的遗传资源。基于此,小编以国务院最新颁发的《国家重点保护野生植物名录》(简称《名录》)为参考,同时结合文献阅读、网站查询等方法,来向大家系统介绍我国濒危植物的基因组研究进展。

图1 本文主要参考书籍
01
国家一级保护野生植物基因组学
研究进展
国务院最新调整的《目录》中,一级保护野生植物共涵盖了蕨类、裸子植物、被子植物及藻类中的54种植物,其中已有基因组发表或报道的物种仅有6种(占比仅有10%),这6种(珙桐、银杏、红豆杉、华山新麦草、貉藻、发菜)植物基因组的研究进展如下。
1.珙桐

文章题目:Genomic analyses of a “living fossil”: The endangered dove-tree
发表期刊:Molecular Ecology Resources
发表时间:2020年1月
测序策略:PacBio+Hi-C
研究结果:珙桐(Davidia involucrata)被称为植物界中的“大熊猫”,这不仅是因为其与大熊猫一样种群濒危,分布地狭窄(仅在中国西南部和中部山区有分布),更是因为其形态与大熊猫一样优雅,深受大众所喜爱。珙桐的两片苞片洁白、硕大,当大风来临时特别像飞舞的鸽子,因此珙桐又名“鸽子树”。本研究中,研究者利用PacBio+Hi-C技术构建了高质量的珙桐基因组,其基因组大小为1169Mb,Contig N50=1.35Mb,珙桐比其近缘种喜树有更多的基因。珙桐与喜树在分化之前经历了一次较近时间的WGD事件,分化后重复序列的扩增对珙桐基因组大小的增加有很大贡献。转录组分析表明,与光合作用相关的基因在珙桐苞片中几乎缺失或表达减少,而与防御和化学相关的基因则大量增加,突出了苞片在保护花朵和吸引传粉者方面的重要作用。重测序分析表明,自第四纪冰期以来珙桐的有效种群大小一直在减小,这表明气候变化可能是珙桐成为濒危的主要原因之一。因此,后续在针对珙桐资源保护时,应充分考虑其对气候的敏感性。
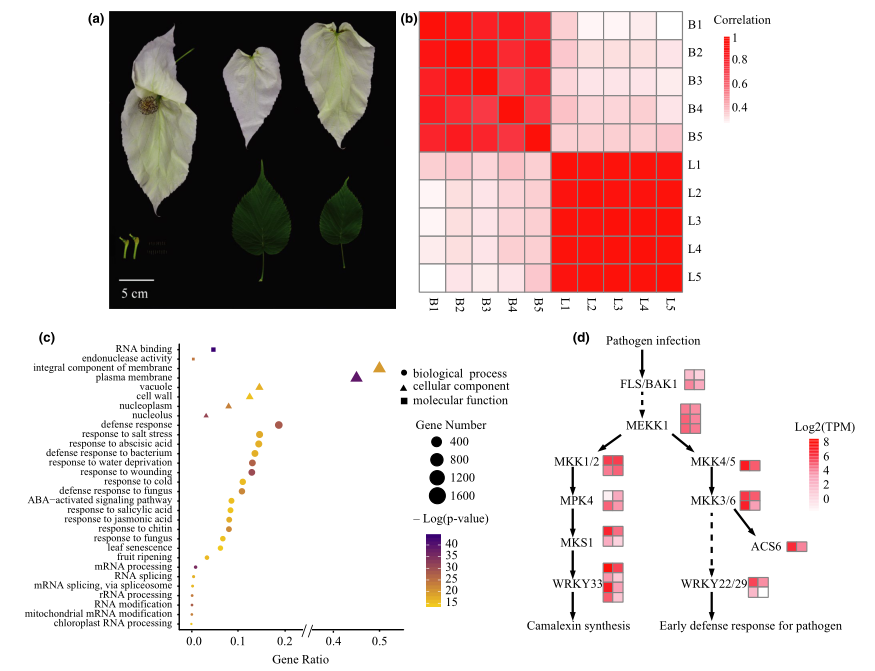
图2 珙桐苞片的转录组分析
2.银杏

文章题目:The nearly complete genome of Ginkgo biloba illuminates gymnosperm evolution
发表期刊:Nature Plants
发表时间:2021年6月
测序策略:PacBio+Hi-C
研究结果:银杏(Ginkgo biloba)作为典型的孑遗物种,是银杏纲现存的唯一成员,不仅具有特殊的进化地位,而且还具有极高的观赏价值和药用价值。研究者组装的银杏基因组大小为9.88Gb(contig N50=1.58Mb),注释得到27832个蛋白编码基因。在研究的植物种类中,银杏基因组内含子的长度是最大的,这进一步说明重复序列不仅促进了基因组的扩展,而且增加了蛋白质编码基因的大小和复杂性。随后研究者利用基因组及转录组来解析银杏特殊的表型,重点关注银杏保留的精子鞭毛、未成形的花、扇形的叶等。总之,本研究对银杏环境适应性、裸子植物进化研究具有重要意义。
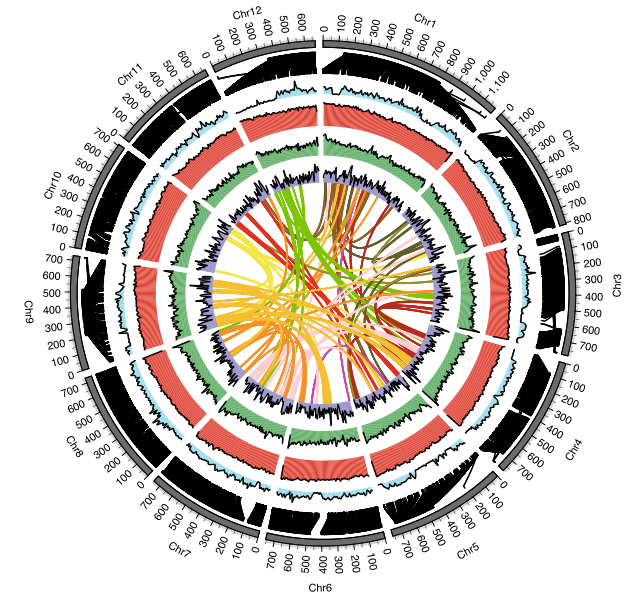
图3 银杏基因组特征
3.红豆杉

文章题目:The Taxus genome provides insights into paclitaxel biosynthesis
发表期刊:Nature Plants
发表时间:2021年7月
测序策略:PacBio+Hi-C
研究结果:红豆杉属于一级濒危保护植物,被众多国家视为“国宝”,其活性成分紫杉醇具有显著的抗癌功效。本研究中,研究者构建了南方红豆杉(Taxuschinensis var. mairei)染色体水平的参考基因组,其基因组大小为10.23 Gb,Contig N50为2.44 Mb。比较基因组分析表明,红豆杉发生了一次WGD事件,重复序列的连续插入使得红豆杉基因组变得庞大;在进化过程中,红豆杉保留了独特的Gypsy和Copia转座子家族和大量紫杉醇合成相关基因。结合基因组、转录组和代谢组数据,研究者系统分析了紫杉醇合成相关基因的基因组定位与协同表达调控,并首次发现了由六个基因串联组成的基因簇与紫杉醇生物合成的前两步密切相关。总之,本研究为紫杉醇生物合成的研究提供了新见解。
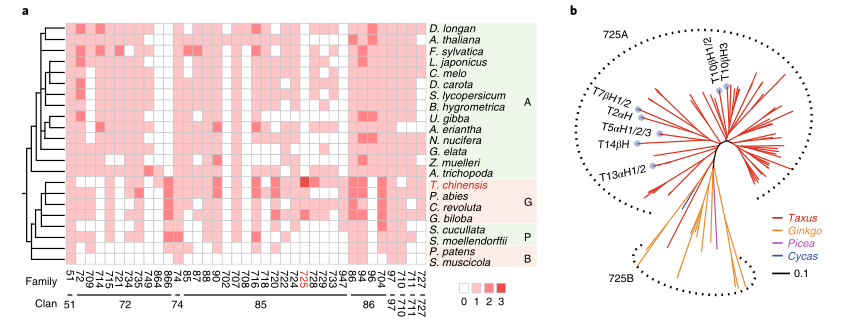
图4 紫杉醇CYP450s的进化及结构特征
4.华山新麦草

华山新麦草(Psathyrostachys huashanica)是我国特有的禾本科植物,属于国家一级保护野生植物,仅在陕西华山的狭小范围内有分布,与同属的其他物种有较大的形态差异和形成间断地理分布。华山新麦草具有耐贫瘠、耐寒、耐旱、优质早熟等品质,是小麦新品种研发的重要资源库。
西北大学生科院李忠虎团队利用PacBio+Hi-C技术构建了华山新麦草的基因组,其基因组大小为6.11Gb,Contig N50=958.8 Kb,将92.3%的序列锚定到7条染色体上。通过注释,发现重复序列占比76.63%,注释得到41842个基因。比较基因组分析表明新麦草中存在449个特有的基因,且与小麦具有很近的亲缘关系。

图5 华山新麦草基因组研究进展
5.貉藻

文章题目:Genomes of the Venus Flytrap and Close Relatives Unveil the Roots of Plant Carnivory
发表期刊:Current Biology
发表时间:2020年6月
测序策略:PacBio+Hi-C
研究结果:貉藻(Aldrovanda vesiculosa)隶属于茅膏菜科、貉藻属浮水草本植物,其仅在黑龙江省有分布,是我国一级濒危保护植物。貉藻可以分泌消化液,来捕捉和消化昆虫,属于“食肉植物”。本研究对捕蝇草(Dionaea muscipula)与其水生姊妹种貉藻(Aldrovanda vesiculosa)和匙叶茅膏菜(Drosera spatulata)进行测序组装,组装的基因组大小为293Mb~3.18Gb,Contig N50 35kb~705kb。转录组分析表明,捕蝇草及其近缘植物显著表达的基因可注释到捕食动物的整个周期活动中(包含猎物吸引、感知、消化和吸收等)。最后结合转录组和ATAC分析,研究者发现捕蝇草、貉藻、匙叶茅膏菜中的WRKY6和WRKY29转录因子对腺体捕食活动相关基因起调控作用。

图6 捕蝇草、貉藻、匙叶茅膏菜的基因组进化
6.发菜

文章题目:Genomic and transcriptomic insights into the survival of the subaerial cyanobacterium Nostoc flagelliforme in arid and exposed habitats
发表期刊:Environmental Microbiology
发表时间:2019年1月
测序策略:PacBio+Illumina
研究结果:发菜(Nostoc flagelliforme)是我国一级保护野生植物,其分布范围主要在我国北部和西北部干旱和荒漠地区,发菜对高强度紫外辐射与极度干旱条件等逆境条件有极强的适应性。本研究中,研究者利用二代和三代测序构建了完整的发菜基因组,其基因组大小为10.23 Mb,包含10825个蛋白编码基因。比较基因组分析表明,与DNA复制、重组和修复相关基因是发菜基因组扩张的主要原因。光合机构保护、单不饱和脂肪酸合成、紫外辐射响应、鸟氨酸代谢相关基因的表达扩增使得发菜可以很好的适应极端环境。转录组分析表明,发菜具有一套独特的复水生活策略,包括PSI组装因子基因的组成型高表达、转录后调控机制参与光合作用的恢复等。总之本研究对蓝藻的逆境适应性研究提供了新见解。
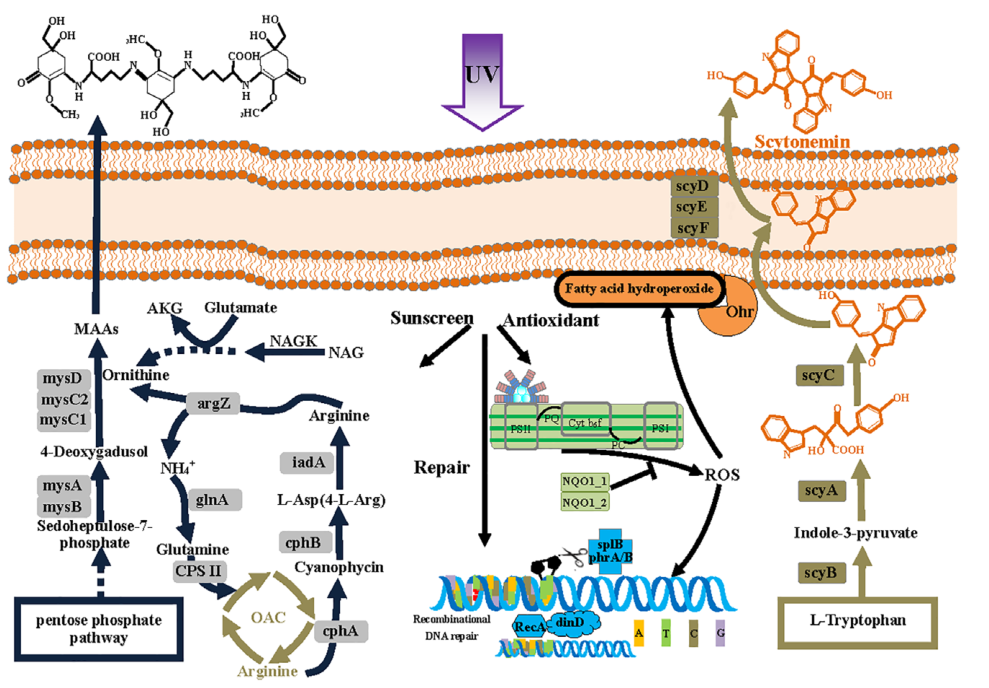
图7 发菜对紫外辐射适应性的示意图
02
国家二级保护野生植物基因组学
研究进展
根据最新《目录》,我国二级保护野生植物共有401种和36类,其中已有基因组发表的物种达几十种,限于篇幅,我们仅选取近两年发表的部分二级保护植物基因组文章进行详细阐述,涉及的物种包括独叶草、闽楠、连香树、铁皮石斛等。
1.独叶草

文章题目:Genome Sequencing of the Endangered Kingdonia uniflora(Circaeasteraceae,Ranunculales) Reveals Potential Mechanisms of Evolutionary Specialization
发表期刊:iScience
发表时间:2020年5月
测序策略:PacBio+Illumina
研究结果:独叶草(Kingdonia uniflora)起源于6700万年前,是我国特有的孑遗植物,其一花一叶的结构以及独特的环境适应性使得独叶草成为被子植物进化研究的模式物种之一。本研究中,研究者构建了独叶草基因组草图,其基因组大小为1.04Gb,包含43301个蛋白编码基因。比较基因组分析发现与DNA修复有关的基因家族在独叶草中发生显著扩张,与环境响应相关的基因家族则发生显著收缩。此外,在独叶草中大部分叶绿体ndh基因都发生了丢失。总之,无性繁殖系统及强大的DNA修复系统共同降低了独叶草的遗传多样性,进而降低了其环境适应性;而相关基因的扩增、收缩也正是独叶草长期适应单一稳定生境的具体表现。
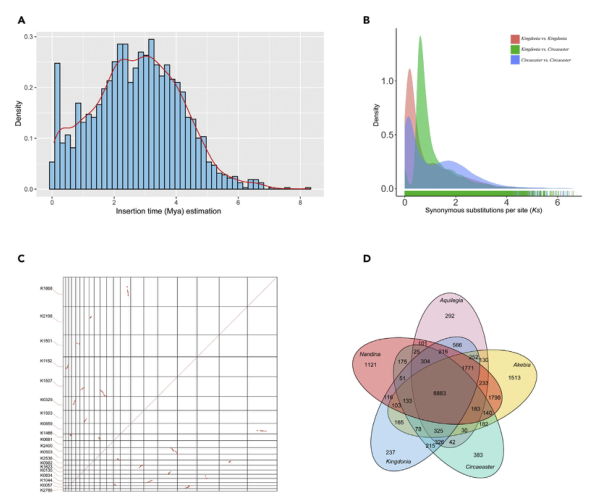
图8 独叶草基因组的进化
2.闽楠

文章题目:The Phoebe genome sheds light on the evolution of magnoliids
发表期刊:Horticulture Research
发表时间:2020年9月
测序策略:PacBio+Illumina
研究结果:闽楠(Phoebe bournei)隶属于木兰类樟目,属于我国二级珍稀保护物种,从古至今以其良好的木材特性而闻名海内外。本研究中,研究者构建的闽楠基因组大小为989.19 Mb,Contig N50=2.05 Mb。比较基因组分析发现闽楠经历了两次WGD事件,且系统进化树支持木兰类与单子叶、双子叶植物互为姐妹枝。研究者在楠木基因组中鉴定了63个MADS-box基因、5个参与光形态发生的FIN219-like基因和2个参与光信号介导茎发育的SAUR50-like基因,这些基因使得闽楠向阳而生,从而形成了笔直的树干。此外,研究者还鉴定了一系列与闽楠木质素生物合成、材质形成相关的基因。总之,本研究为木兰类植物进化、闽楠的多样性和保护提供了新见解。
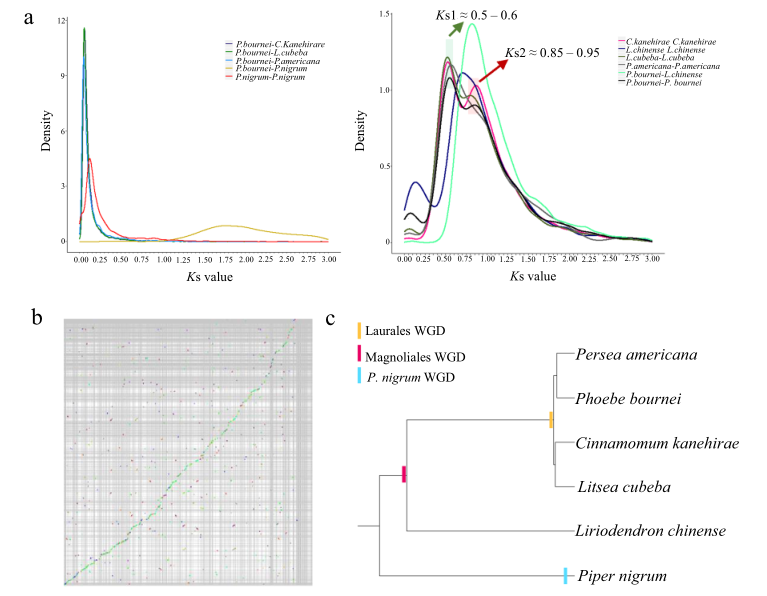
图9 闽楠基因组的WGD分析
3.连香树

文章题目:Genomic insights on the contribution of balancing selection and local adaptation to the long-term survival of a widespread living fossil tree, Cercidiphyllum japonicum
发表期刊:New Phytologist
发表时间:2020年7月
测序策略:PacBio+Illumina+Hi-C+ 10× Genomics
研究结果:连香树(Cercidiphyllum japonicum)隶属于连香树科连香树属,是东亚特有的第三纪孑遗落叶林,同时也是珍稀濒危变色树种。连香树在多种气候带中都有分布,是研究第三纪孑物种演化和适应性的模式材料。在本研究中,研究者利用多种测序技术构建了高质量的连香树基因组,其基因组大小为719Mb,Contig N50=494.88Kb,注释得到27393个基因。随后结合比较基因组、99个个体的重测序分析,研究者从基因组层面构建了连香树属的物种演化历史,解析了连香树广泛环境适应性的遗传机制,从而为连香树的进化及种群保护提供了新的见解。
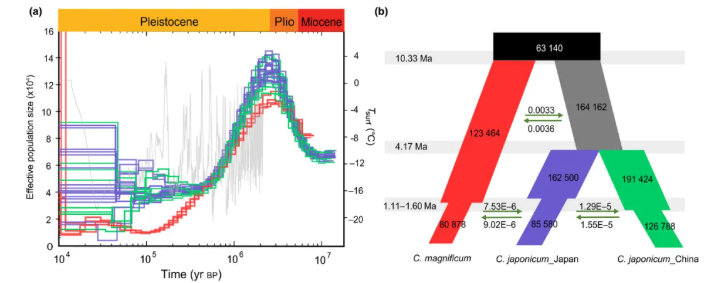
图10 连香树的有效群体大小和种群历史动态分析
4.铁皮石斛

文章题目:The chromosome-level reference genome assembly for Dendrobium officinaleand its utility of functional genomics research and molecular breeding study
发表期刊:Acta Pharmaceutica Sinica B
发表时间:2021年7月
测序策略:PacBio+Illumina+Hi-C
研究结果:铁皮石斛(Dendrobium officinale)隶属于兰科石斛属,是我国特有的名贵中药材,其具有养阴生津、润肺明目、抗癌防老等杰出功效,被誉为中华“九大仙草”之首,且已被列入《中华人民共和国药典》与药食同源目录。但由于缺乏高质量的参考基因组,近年来铁皮石斛功能基因组与分子育种研究进展较慢。本研究中,研究者构建了首个染色体水平的铁皮石斛基因组,结合比较基因组、转录组、化学成分分析等方法对铁皮石斛重要次生代谢产物的关键酶基因进行了挖掘,并通过GWAS分析初步确定了与铁皮石斛产量相关的基因,为铁皮石斛功能基因组学和分子育种研究提供了新见解。
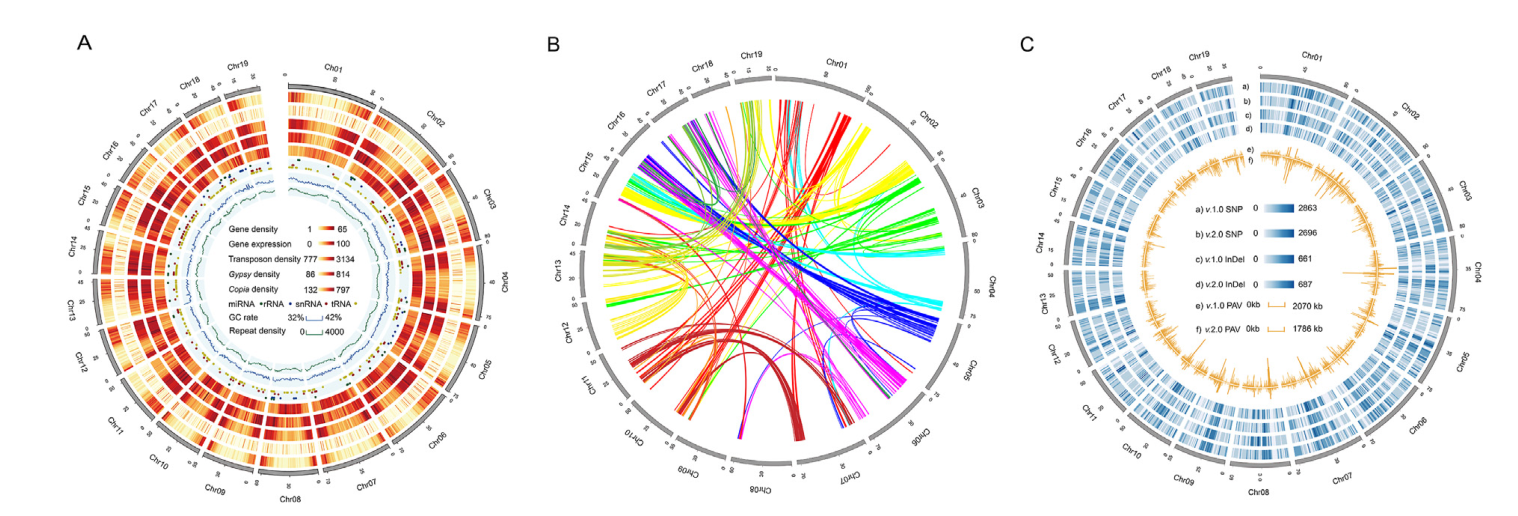
图11 铁皮石斛基因组特征
最后,小编将《重点保护野生植物名录》、濒危植物基因组研究文献及思路进行了打包整理。关注“永利集团3044”公众号,转发此篇公众号文章到朋友圈,然后在微信公众号后台发送截图信息,即可免费领取!
参考文献:
[1] Chen Y, Ma T, Zhang L, et al. Genomic analyses of a “living fossil”: The endangered dove‐tree[J]. Molecular ecology resources, 2020, 20(3): 756-769.
[2] Liu H, Wang X, Wang G, et al. The nearly complete genome of Ginkgo biloba illuminates gymnosperm evolution[J]. Nature Plants, 2021, 7(6): 748-756.
[3] Xiong, X., Gou, J., Liao, Q. et al. The Taxus genome provides insights into paclitaxel biosynthesis. Nat. Plants 7, 1026–1036 (2021).
[4] 李忠虎. 华山新麦草基因组测序研究进展[C]// 第十届全国小麦基因组学及分子育种大会. 2019.
[5] Palfalvi G, Hackl T, Terhoeven N, et al. Genomes of the venus flytrap and close relatives unveil the roots of plant carnivory[J]. Current Biology, 2020, 30(12): 2312-2320. e5.
[6] Shang J L, Chen M, Hou S, et al. Genomic and transcriptomic insights into the survival of the subaerial cyanobacterium Nostoc flagelliforme in arid and exposed habitats[J]. Environmental microbiology, 2019, 21(2): 845-863.
[7] Sun Y, Deng T, Zhang A, et al. Genome sequencing of the endangered Kingdonia uniflora (Circaeasteraceae, Ranunculales) reveals potential mechanisms of evolutionary specialization[J]. Iscience, 2020, 23(5): 101124.
[8] Chen S P, Sun W H, Xiong Y F, et al. The Phoebe genome sheds light on the evolution of magnoliids[J]. Horticulture research, 2020, 7(1): 1-13.
[9] Zhu S, Chen J, Zhao J, et al. Genomic insights on the contribution of balancing selection and local adaptation to the long‐term survival of a widespread living fossil tree, Cercidiphyllum japonicum[J]. New Phytologist, 2020, 228(5): 1674-1689.
[10] Niu Z, Zhu F, Fan Y, et al. The chromosome-level reference genome assembly for Dendrobium officinale and its utility of functional genomics research and molecular breeding study[J]. Acta Pharmaceutica Sinica B, 2021.
 微信公众号
微信公众号


 027-87224696
|
027-87224696
| marketing@frasergen.com
|
marketing@frasergen.com
|
